

Revisión
Enfermedad de Tarui: revisión y perspectivas bioinformáticas
Tarui´s disease: Review and bioinformatic perspectives
Actual. Med. 2016; 101: (798): 101-108 DOI: 10.15568/am.2016.798.re01
Enviado: 31-03-2016
Revisado: 11-07-2016
Aceptado: 20-07-2016
RESUMEN
La enfermedad de Tarui es una enfermedad autosómica recesiva ocasionada por una deficiencia de la actividad fosfofructoquinasa (FFQ) en el músculo esquelético, una enzima clave de la glicólisis. Dicha alteración tiene como consecuencia directa la imposibilidad de producir energía a partir del consumo de la glucosa circulante en sangre o la que se almacena en forma de glucógeno. Debido a esto, clásicamente los síntomas incluyen intolerancia al ejercicio, mialgias, debilidad y fatiga, pero, pese a que se han detectado al menos 22 mutaciones del gen que codifica por la enzima, pacientes con las mismas variantes han mostrado diferente sintomatología, lo cual establece una relación genotipo-fenotipo poco clara. Mecanismos regulatorios, y no solamente la enzima misma, pueden explicar la modulación de la actividad FFQ y con ello la diversidad evidenciada con los cerca de 100 casos reportados con la enfermedad de Tarui. El diagnóstico definitivo se obtiene de la evaluación de biopsia de músculo, pruebas inmunohistoquímicas, medición de la actividad de la enzima y prueba de glucosa exógena previo a ejercicio. Actualmente no existe un tratamiento específico y normalmente se brinda terapia específica para los síntomas. Nuevas perspectivas con el uso de herramientas bioinformáticas tienen el potencial de estudiar la enzima FFQ con análisis estructurales de la molécula o bien, los diversos sistemas biológicos que la regulan.
Palabras clave: Enfermedad de Tarui, Glucogenosis tipo VII, miopatía metabólica, fosfofructoquinasa.
ABSTRACT
Tarui’s disease is an autosomal recessive disorder caused by a deficiency of phosphofructokinase activity (FFQ) in skeletal muscle, a key enzyme of glycolysis. This alteration has a direct consequence of the impossibility of producing energy from the consumption of circulating-blood glucose or stored as glycogen. Because of this, classically symptoms include exercise intolerance, muscle pain, weakness and fatigue, but despite that have been detected at least 22 mutations of the gene coding for the enzyme, patients with the same variants have shown different symptoms, which establishes a genotype-phenotype relationship unclear. Regulatory mechanisms, and not only the enzyme itself, can explain the modulation of the FFQ activity and thus the diversity evidenced with about 100 cases reported Tarui’s disease. The definitive diagnosis is obtained from the evaluation of muscle biopsy, immunohistochemistry, measurement of enzyme activity and exogenous glucose test prior to exercise. Currently there is no specific treatment and therapy usually is given for symptoms. New perspectives using bioinformatic tools have the potential to study the FFQ enzyme with structural analysis of the molecule or the various biological systems that regulate it.
Keywords: Tarui´s disease, glycogenosis type VII, metabolic myopathy, phosphofructokinase.
Leer Artículo Completo
INTRODUCCIÓN
El monosacárido de glucosa es la fuente primaria de producción de energía de las células y que puede ser almacenado en forma de polímeros de glucógeno, esencialmente en el hígado y el músculo esquelético. Diferentes mecanismos en redes bioquímicas describen el balance entre la glucosa disponible para producción de energía o para su almacenamiento. Cuando es requerida, por ejemplo en el ayuno o durante el ejercicio, la glucosa puede ser obtenida de forma directa de la ingesta de alimentos o de la degradación del glucógeno (glucogenólisis) a glucosa-1-fosfato. La producción de energía se da a través de la glicólisis, en la que la glucosa es degradada hasta producir ATP (adenosina trifosfato), la moneda energética, además de piruvato y NADH (dinucleótido de nicotinamida y adenina) que pueden continuar a ciclo de Krebs o a respiración anaerobia. Cuando las necesidades energéticas han sido satisfechas, diversos mecanismos regulatorios permiten el almacenamiento del exceso de glucosa en forma de glucógeno, en un proceso llamado gluconeogénesis (1,2).
La gluconeogénesis, la glucogenólisis y la glucólisis son procesos celulares que son mediados por enzimas y altamente regulado, tanto por los mismos carbohidratos como por otros tipos de moléculas. La deficiencia parcial o total de la actividad enzimática en las reacciones de dichos procesos se constituyen como un grupo de trastornos del metabolismo llamados enfermedades del almacenamiento del glucógeno (EAG) (2). Si la afectación de estos procesos es de forma directa en las enzimas, la enfermedad se denomina enfermedad primaria, mientras que si la alteración es por medio de reguladores de su metabolismo, entonces se llaman enfermedades secundarias (3).
ENFERMEDAD DE TARUI
Enfermedades del almacenamiento del glucógeno
Las enfermedades primarias del almacenamiento del glucógeno, también conocidas como glucogenosis, corresponden a una categoría única de trastornos metabólicos hereditarios que se describieron inicialmente a principios del siglo XX. Fueron uno de los primeros errores innatos del metabolismo con deficiencia enzimática en ser identificados y que afectan la homeostasis energética del músculo esquelético, corazón, hígado y, con menor frecuencia, el sistema nervioso central (1). Usualmente, presentan un patrón de herencia autosómico recesivo y su incidencia se estima en 1 caso por cada 20,000-43,000 nacimientos vivos (4,5).
Dadas las variaciones que existen en la edad de inicio, los síntomas, la morbilidad y la mortalidad de las EAG, la clasificación de estos trastornos ha sido motivo de discusión constante (3). Sin embargo, las propuestas clásicas se han enfocado en realizar la agrupación de acuerdo a la deficiencia enzimática y, en la medida de lo posible, el patrón clínico. Se han identificado al menos 14 tipos de glucogenosis (4), cuyos efectos primordialmente se ven reflejados en los tejidos hepático y muscular esquelético (6), y en menor grado miocardio, sistema nervioso central y riñones (7).
En la tabla 1 se muestra la clasificación de las EAG, según la enzima afectada y el compromiso hepático o muscular. Como se mencionó previamente, algunos casos específicos pueden afectar bazo, corazón y otros órganos (4). Por ejemplo, cuando hay ausencia de actividad maltasa ácida en la enfermedad de Pompe (Tipo II) hay afectación de casi todos los órganos, incluyendo al corazón (2,4).
Respecto a la incidencia de las EAG según su clasificación, la tipo I es la más frecuente, con el 90% de las EAG. De las que afectan tejido muscular, la enfermedad de Tarui es la segunda más frecuente, solo superada en frecuencia por la enfermedad de McArdle (3). Los principales síntomas de las EAG que afectan músculo incluyen intolerancia al ejercicio, debilidad muscular, mialgias, calambres y mioglobinuria, y su aparición puede variar tanto en edad como en intensidad. Así, el pronóstico de cada paciente con una EAG dependerá de la enzima que se encuentre afectada y el tipo de alteración que tiene, aunque es común que en las presentaciones infantiles los pacientes fallezcan en el transcurso de los primeros años de vida; por el contrario, en otros pacientes la presentación puede ser tardía o no dar otros síntomas aparte de la intolerancia al ejercicio (7).
Aspectos generales de la enfermedad de Tarui
La enfermedad de Tarui es el tipo VII de las glucogenosis. Tiene consecuencias predominantemente en músculo esquelético debido a una deficiencia en la actividad FFQ celular. La FFQ constituye una enzima clave en el proceso de glicólisis, ya que se encarga de la conversión de fructosa-6-fosfato a fructosa-1,6-bifosfato en forma irreversible (8). De esta manera, en la enfermedad de Tarui hay un aporte comprometido de energía a partir de la glucosa, lo cual es crítico en condiciones como el ejercicio.
Estructuralmente, la FFQ es un tetrámero que derivada de tres loci genéticamente distintos y que codifica por isoformas de músculo, hígado, y plaquetas. En músculo y el hígado, las FFQs son homotetrámero de 4 subunidades M y 4 subunidades L, respectivamente, mientras que los eritrocitos contienen tanto subunidades L y M (3). En el caso de las plaquetas, las subunidades son tipo P, aunque también se conoce como tipo F (de fibroblastos). Los genes de las subunidades tipos M, L y P se ubican en los cromosomas humanos 12, 21 y 10, respectivamente (9).
Históricamente, la primera descripción de esta patología la realizaron Tarui y colaboradores (1965), en tres parientes japoneses (una mujer de 20 años de edad y dos hombres de 23 y 27 años de edad), cuyos padres eran primos. Sus manifestaciones clínicas fueron descritas como facilidad para fatigarse, debilidad y rigidez muscular posterior a ejercicios intensos o prolongados, sin elevación de lactato en sangre durante el test de isquemia y uno de ellos con mioglobinuria post-ejercicio (1,0). En los años posteriores y hasta la actualidad, los reportes alcanzan al menos 100 casos de pacientes con esta enfermedad, con una incidencia particular en personas con ascendencia judía Ashkenazi, en las zonas de Europa, Japón y Estados Unidos (7,8,1,1). En niños, la ocurrencia de la enfermedad de Tarui fue de al menos una docena de casos entre 1987 y 2008 (1,2).
En los años más recientes, se destacan dos reportes de los años 2012 y 2015. El caso reportado en 2012, una mujer marroquí de 41 años de edad con deficiencia de FFQ presentó debilidad muscular lentamente progresiva desde la infancia, sin episodios de rabdomiólisis o anemia hemolítica, pero la exploración física mostró una leve atrofia de los muslos. La biopsia de músculo reveló almacenamiento masivo de glucógeno en la mayoría de las fibras musculares y una actividad FFQ en el músculo totalmente ausente (1,3). Asimismo, Auranen y colaboradores (2015) reportaron el caso de dos hermanos con la enfermedad de Tarui con manifestaciones clínicas clásicas y formación de acúmulos de poliglucosanos y una mezcla compleja de proteínas en la biopsia de músculo (1,4).
Fisiopatología
En la enfermedad de Tarui, la deficiencia de la actividad FFQ tiene como consecuencia inmediata el bloqueo de la glicólisis. Esta ruta metabólica es la encargada de oxidar la glucosa hasta piruvato, con el objetivo de obtener energía para la célula. El catabolismo de glucosa consiste de 10 reacciones enzimáticas acopladas que permiten la obtención de ATP, NADH y dos moléculas de piruvato, proceso que ocurre en el citoplasma celular. En dependencia de las condiciones de oxigenación, el piruvato puede dirigirse a diversas rutas metabólicas para producir más energía (1).
Las reacciones que componen la glicólisis se agrupan en 2 fases metabólicas, una primera de gasto de energía y otra de producción de energía. En la primera fase, la reacción inicial consume una molécula de ATP para fosforilar la glucosa a glucosa-6-fosfato para luego convertirla en fructuosa-6-fosfato. Una segunda molécula de ATP se requiere para el paso de una segunda fosforilación de la molécula da origen la fructuosa-1,6-bifosfato, gracias a la enzima FFQ. Otras dos reacciones terminarían de originar 2 moléculas de gliceraldehído-3-fosfato, producto final de la fase de gasto de energía (1,5).
En la fase de producción de energía, la oxidación de cada gliceraldehído-3-fosfato permite la producción de NADH a partir del NAD+, y posteriormente la producción de 2 moléculas de ATP y el piruvato, con la acción final de la piruvato quinasa. Así, el balance entre consumo y producción, se obtiene un balance a favor energético de 2 ATP y 2 NADH, así como 2 moléculas de piruvato (1).
En la respiración celular aerobia, que ocurre en mitocondria, los electrones del NADH son cedidos a la cadena de transporte de electrones, los cuales alteran el potencial de membrana al mover H+ al espacio intermembrana de la mitocondria, y su paso de regreso por el complejo ATP-sintasa logra la producción de más ATP, además de la producción de ATP y NADH que se obtiene del piruvato que ingresa a ciclo de Krebs. En respiración celular anaerobia, procesos de fermentación pueden obtener alternativamente energía del piruvato, con la producción de lactato gracias a la deshidrogenasa láctica. El transporte de lactato desde el músculo hacia hígado puede hacer que se reconvierta a piruvato y luego a glucosa, la cual puede ser llevada de nuevo a músculo como fuente de energía (proceso conocido como ciclo de Cori).
En la enfermedad de Tarui, la fase inicial de la glicólisis arranca hasta la producción de fructuosa-6-fosfato, con el consumo de ATP, pero la actividad alterada de la FFQ impide que se continúe hasta la fase de producción de energía, ni la producción de piruvato ni NADH, lo cual aún compromete más la continuación de procesos como la respiración celular (condiciones aeróbicas) o fermentación (condiciones anaerobias) para obtener más energía. De esta forma, la deficiencia de la FFQ altera por completo la disponibilidad de energía de las células por la caída en la producción de ATP, piruvato y poder reductor en forma de NADH (16).
Además, dado que los carbohidratos y los ácidos grasos son la principal fuente de energía para la contracción y relajación muscular, la deficiencia enzimática imposibilita que el glucógeno almacenado en el músculo o bien la glucosa circulante en sangre puedan ser usadas como fuentes de energía, por lo que la vía metabólica se desplaza hacia la lipólisis (17). Sin embargo, esta vía no logra compensar de forma equivalente a la glicólisis, lo que compromete la producción de energía suficiente durante el ejercicio físico. El inadecuado funcionamiento de la FFQ también conlleva al depósito o almacenamiento excesivo de glucógeno intracelular, lo cual provoca compresión y desplazamiento de la maquinaria contráctil de las fibras musculares, con las manifestaciones clásicas de la enfermedad a nivel muscular (18). Sin embargo, la FFQ no solamente es de relevancia en esta enfermedad metabólica, sino que en conjunto con otras enzimas, ha sido estudiada en otros tipos de padecimientos como lo son la obesidad, diabetes y cáncer (19), por lo que su deficiencia también puede inducir padecimientos paralelos en los pacientes.
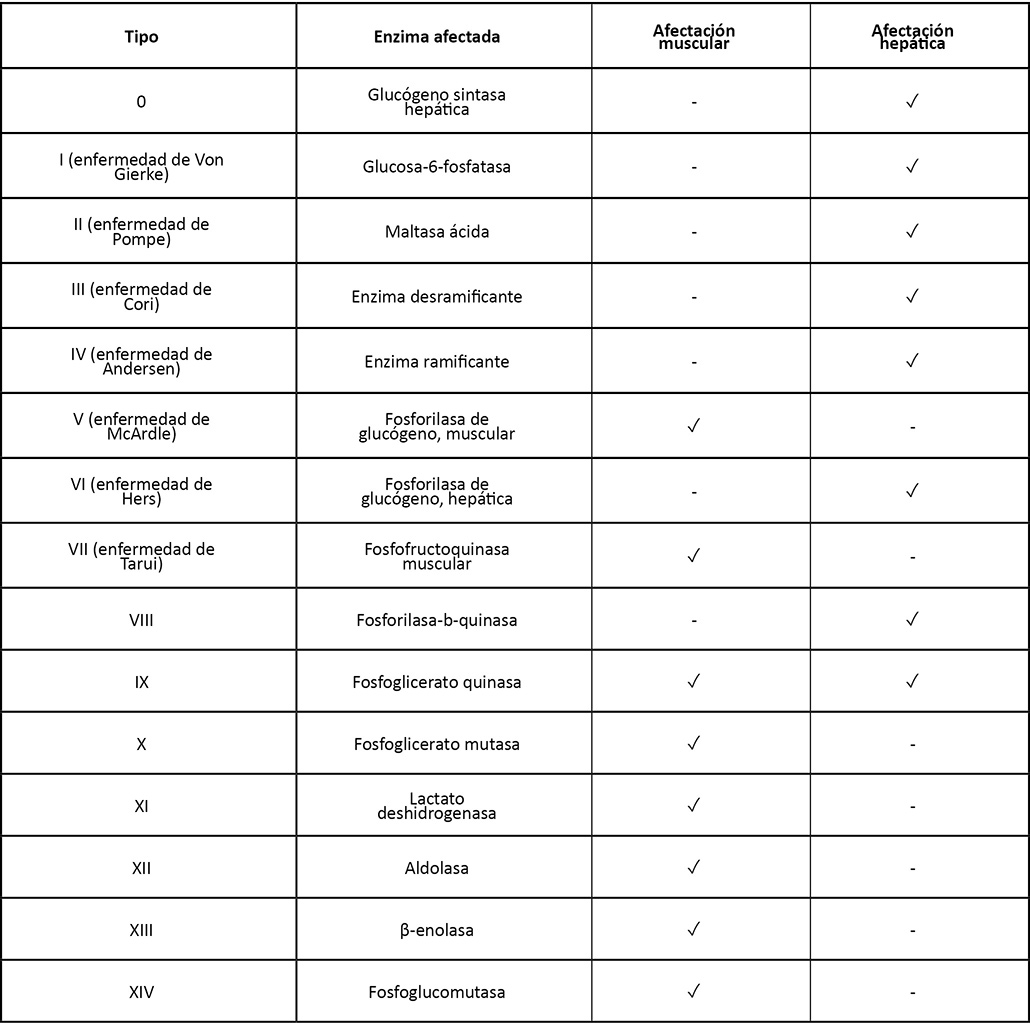
Tabla 1 . Clasificación de las enfermedades del almacenamiento del glucógeno.
Fuente: Elaboración propia.
A nivel sanguíneo, la glicólisis deficiente muestra, típicamente, cerca de un 50% de actividad enzimática FFQ, respecto a pacientes sin la enfermedad. Esta reducción del flujo glicolítico puede limitar gravemente el suministro de energía y resultar en hemólisis, comúnmente reportada en la enfermedad de Tarui (3,2,0). Entre 2001 y 2003, Ronquist y Waldenstro (2001) observaron que los eritrocitos de las personas con la enfermedad de Tarui presentaban niveles elevados de calcio intraeritrocitario. Mostraron que en la enfermedad se produce un aumento de la hidrólisis de ATP con una acelerada extrusión del calcio ATP-dependiente, sugiriendo un aumento de la permeabilidad de calcio en la membrana celular. Los mismos autores exponen que este fenómeno ocurre de forma paralela a la deficiencia de la FFQ y que, a nivel sanguíneo, explicaría la rigidez de la membrana y hemólisis de las células del sistema reticuloendotelial, mientras que en músculo llevaría a fatiga debido a la regulación disfuncional del calcio, posiblemente más que la deficiencia misma de la enzima (11,20).
Manifestaciones clínicas
Los pacientes diagnosticados con la enfermedad de Tarui pueden presentar miopatías y hemólisis, ya sea como condiciones únicas o en combinación, o bien podrían cursar asintomáticos (8,21). La miopatía se manifiesta como un conjunto de síntomas que abarcan desde debilidad leve hasta intolerancia marcada al ejercicio, calambres musculares, disminución de reflejos tendinosos y niveles séricos elevados de enzimas musculares (21). No obstante, en algunas personas la evolución y síntomas son atípicos pese a que se detecten las mismas variantes de la enzima, lo cual se traduce en una heterogeneidad fenotípica (Nakajima, Raben, Hamaguchi, & Yamasaki, 2002). Debido a esto, se han propuesto al menos cuatro presentaciones clínicas de la enfermedad de Tarui, resumidas en la tabla 2.
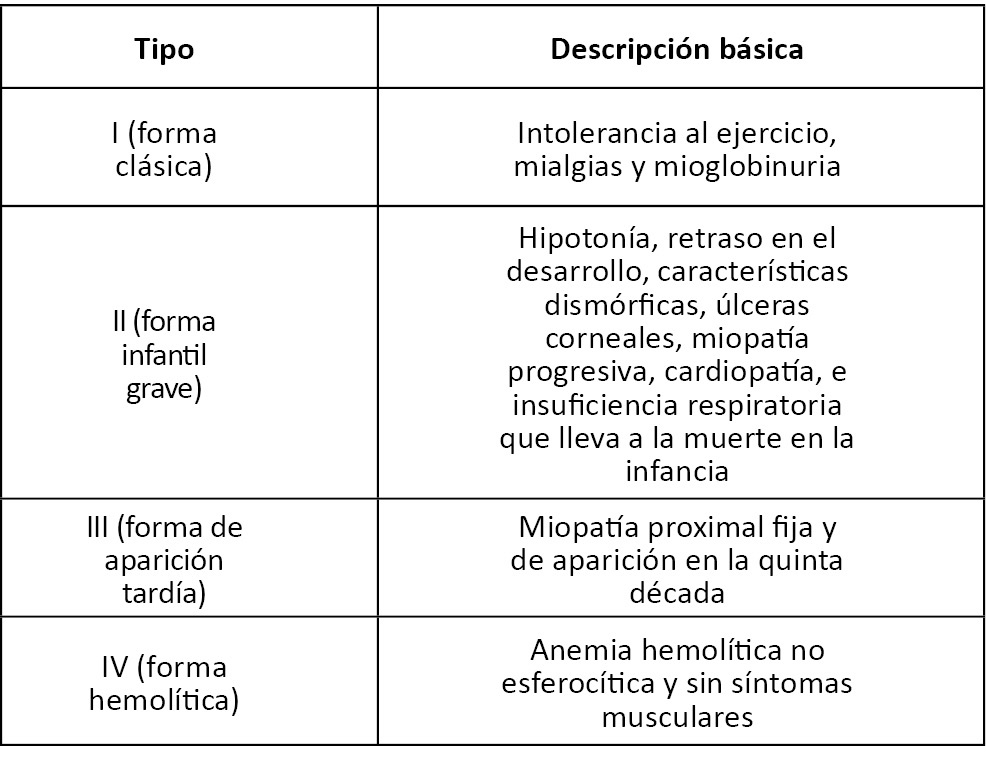
Tabla 2. Presentaciones clínicas de la enfermedad de Tarui
Fuente: Elaboración propia.
Algunos pocos casos reportados han mostrado alteraciones del crecimiento y manifestaciones con una desregulación del metabolismo del ácido úrico, así como afectación del sistema nervioso central y del corazón (5,23). Finsterer y colaboradores (2002) describieron el caso de una paciente con enfermedad de Tarui que desarrolló epilepsia focal (posiblemente consecuencia de acúmulo de glucógeno en cerebro) y síntomas clásicos a nivel muscular, disminución de reflejos tendinosos, test anormal de isquemia en antebrazo, electromiograma con compromiso miogénico y biopsia muscular anormal. Adicionalmente, la paciente presentaba compromiso cardiaco, que se manifestó inicialmente por dolor torácico y luego con un electrocardiograma que mostró bajo voltaje (atribuido al depósito de glucógeno). Además, se evidenció un engrosamiento del miocardio sin asociación con ninguna cardiopatía, por lo cual dicho hallazgo fue atribuido también al depósito de glucógeno (21). Finsterer y Stöllberger (2008) dieron seguimiento a esta paciente, a quien se le detectó un engrosamiento progresivo de las válvulas cardiacas como consecuencia del depósito de glucógeno, lo cual constituyó el primer caso clínico reportado en la literatura de dicha alteración en una persona con enfermedad de Tarui (23).
Diagnóstico
Dado que la afectación muscular esquelética en la enfermedad de Tarui se caracteriza por síntomas poco específicos, sin evidencias francas de la enfermedad ni diferenciales de otros padecimientos, el diagnóstico de esta miopatía metabólica puede ser difícil. Dependiendo de la presentación clínica y de la intensidad con la que ocurra, los pacientes pueden no presentar hallazgos objetivos y claros en el examen neurológico, electromiografía y exámenes de rutina de laboratorio, los cuales con cierta frecuencia se describen como normales (4). En muchos casos, la sospecha de la enfermedad de Tarui se da por exclusión de otros padecimientos, y pese a que los síntomas podrían ser leves e inespecíficos el diagnóstico correcto es importante para prevenir posibles complicaciones graves, como mioglobinuria y fallo renal (4,24).
Las primeras sospechas suelen provenir de las crisis agudas reversibles, que se manifiestan principalmente en forma de fatiga prematura y contracturas musculares, con frecuencia acompañadas de rabdomiolisis con elevada creatina quinasa en suero y, en ocasiones, por mioglobinuria (2). También es posible observar ictericia, niveles elevados de creatina quinasa (CK), hiperuricemia, reticulocitosis y aumento de la bilirrubina sérica. La forma de aparición tardía se presenta con calambres y mialgias en la edad adulta, aunque la capacidad de ejercicio ya es baja desde la infancia (3,9). Los pacientes con la forma infantil comúnmente mueren durante el primer año de vida y podrían mostrar signos de artrogriposis y retraso mental (9).
Ante las sospechas clínicas, el diagnóstico de la glucogenosis de tipo VII se basa especialmente en la biopsia del músculo, que muestra la acumulación de glucógeno asociado con la deficiencia de FFQ por histoquímica (3). Las biopsias musculares a menudo muestran vacuolización interna con el almacenamiento de glucógeno, que pueden ser reveladas por tinción de PAS evidenciando el depósito en el sub-sarcolema y áreas intra-miofibrilares, aunque en algunos casos los aspectos morfológicos podrían ser casi normales (3,25). Además, también se pueden tener hallazgos de valores de glucosa-1- fosfato, glucosa-6-fosfato, y la fructosa-6-fosfato altos en el tejido, mientras que se reducen la fructosa-1,6-bifosfato y triosa-fosfato (7),(9). La presencia de depósitos de poliglucosanos se ha informado en algunos casos de deficiencia de FFQ, especialmente en pacientes de edad avanzada y que podría ser secundaria al acúmulo de glucosa-6-fosfato, activador de la sintasa de glucógeno. Esta característica de acumulación de poliglucosanos es característica típica de la deficiencia de la enzima de ramificación de la EAG tipo IV, por lo que debe también realizarse diagnóstico diferencial al respecto (13).
Las pruebas para evaluar la presencia de FFQ por histoquímica suelen ser negativas y tradicionalmente ha sido un factor diagnóstico clave para explicar la enfermedad (2). Sin embargo, recientemente se reportó el caso de dos hermanos con la enfermedad de Tarui donde las pruebas inmunohistoquímicas mostraron cantidades normales de la FFQ, y la mutación identificada no redujo la cantidad de enzima en general, pero sí reducía severamente su actividad enzimática. Los resultados indicaron que la histoquímica convencional utilizada anteriormente como “estándar de oro” para el diagnóstico de esta glucogenosis debe interpretarse con cautela y es requerido realizar las mediciones de actividad enzimática, además de pruebas de genética molecular, como las modalidades de pruebas definitivas para el diagnóstico (14).
Dado que algunas otras glucogenosis que afectan músculo podrían tener síntomas y hallazgos de laboratorio similares, la prueba de ingesta de glucosa o sacarosa, o bien la inyección de glucagón previo al ejercicio, es de carácter diferencial., particularmente de la enfermedad de McArdle. Ante la administración del azúcar antes del ejercicio muscular, los síntomas de los pacientes con enfermedad de Tarui muestran que los síntomas se exacerban (Haller & Vissing, 2004), pero la administración exógena de azúcar permite mejorar la tolerancia al ejercicio en pacientes con enfermedad de McArdle (22, 27). Además, con un mecanismo similar a la administración de azúcar, según refieren Haller y Lewis (1991), las comidas altas en carbohidratos pueden exacerbar la intolerancia al ejercicio en los pacientes con enfermedad de Tarui, por una parte porque su condición les impide utilizar la glucosa o el glucógeno en la producción de ATP, y por otra parte porque el aumento de la glicemia en sangre bloquea la disponibilidad de ácidos grasos libres y cetonas que el músculo podría utilizar como fuente de energía (Haller & Lewis, 1991). A dicho bloqueo en el uso de esa fuente alternativa de combustible energético, común e indispensable en los pacientes con enfermedad de Tarui, se le conoce como fenómeno out of wind (17).
Además, existen pruebas de secuenciación genética para las glucogenosis, que pueden ser aplicadas a pacientes cuyos padres o familiares ya han sido diagnosticados con alguna de estas enfermedades. Las muestras pueden ser obtenidas de vellosidades coriónicas o líquido amniótico (7).
Tratamiento
Actualmente no existe un tratamiento específico para la enfermedad de Tarui, pero entre las recomendaciones generales para estos pacientes se encuentran evitar ejercicio físico de alta intensidad (5), así como una dieta alta en aminoácidos de cadena ramificada y suplementación con vitamina B6, con el fin de promover reparación muscular (7). A diferencia de otras glucogenosis, para las cuales se han desarrollado terapias génicas (por ejemplo, enfermedad de Pompe o glucogenosis tipo II), en el caso de la enfermedad de Tarui el tratamiento continúa siendo paliativo (29).
Aspectos genéticos y moleculares asociados a la enfermedad
Como fue mencionado anteriormente, existen tres isoformas de la FFQ humana en músculo, hígado, y plaquetas, codificadas por los genes pkf-m en el cromosoma 12, pfk-l en el cromosoma 21 y pfk-p en el cromosoma 10, respectivamente (9). El gen pfk-m humano es un gen de copia única que contiene aproximadamente 30 kb de ADN genómico y 24 exones (30).
Las primeras evidencias de que las mutaciones de la isoforma muscular de la FFQ era causales de la enfermedad de Tarui provino de estudios de Western Blot, mostrando bandas inmunorreactivas a pesar de la pérdida de la actividad enzimática en cultivo de células de músculo de pacientes con la enfermedad (31).
Fue hasta 1990 que la primera mutación en el gen pfk-m fue descrita, con un patrón de herencia autosómica recesiva y en uno de los tres pacientes que fueron reportados con la enfermedad, en 1965, provocando una deleción en el marco de 75 pb que se encuentra en un dominio probable del sitio de activación de la enzima con AMP/ADP (Nakajima et al., 1990).
Desde entonces, se han reportado al menos 22 mutaciones, incluidas mutaciones sin sentido, mutaciones silentes y mutaciones de corte y empalme (Brüser, Kirchberger, & Schöneberg, 2012; Musumeci et al., 2012; Nakajima et al., 2002; Nichols et al., 1996; Sherman et al., 1994; Tsujino et al., 1994; Vives-Corrons, Koralkova, Grau, Mañú Pereira, & Van Wijk, 2013). Sin embargo, aunque muchos de los pacientes con las mutaciones descritas tienen síntomas clásicos de la enfermedad, la gravedad y manifestaciones variadas son comunes, por lo que no se ha reconocido una clara correlación genotipo-fenotipo en la enfermedad de Tarui (13).
De acuerdo con Raben y Sherman (1995), la mutación más frecuente es un defecto en el sitio de empalme 5’ del intrón 5, resultando en una deleción en el marco de la secuencia de exón 5, que en su momento representaba el 68% de los alelos mutantes de personas judías Ashkenazi, mientras que, en la misma población, la deleción de una base en el exón 22, representaba alrededor del 27% de las mutaciones (36).
Otras de las mutaciones identificadas corresponde a un caso de tres hermanos de una familia sueca con sintomatología muy distinta y en la que, para dos de ellos, se presentó un cambio de base en el último nucleótido del exón 13 (G1127A), resultando una retención parcial del intrón 13. Además, en otro caso se identificó un cambio de una base en el intrón 16 (G64A), resultando en la retención parcial del intrón 16 (24)
Más recientemente, en 2012, se reportaron 5 mutaciones no detectadas previamente. La primera, una mutación intrónica homocigota (IVS6-2 A>C), reveló una activación de dos sitios de empalme críticos en el exón 7, dando lugar a dos especies de ARN mensajeros, uno con una deleción de 5 pb y el otro con una deleción de 12 pb. Esta segunda variante de ARNm condujo a una deleción en marco de cuatro aminoácidos en el péptido. Además, otra mutación intrónica (IVS5-3 A>G) generó la inserción de AG entre el exón 5 y el exón 6, que condujo a un desplazamiento del marco de lectura y una terminación prematura del trascripto. También se describieron tres nuevas mutaciones sin sentido (D591A, S180C, G57V) y que involucró residuos altamente conservados y cuyas predicciones en modelos estructurales indican efectos negativos sobre la estructura o función de la proteína (9).
En 2015, se reportó el caso de dos hermanos con la enfermedad de Tarui, en la que se detectó el cambio homocigoto, p.R39Q, en el exón 4. Anteriormente, diferentes mutaciones que afectan a la misma posición habían sido descritas con un cambio de arginina a prolina en estado homocigoto, o alanina en heterocigoto. Al realizar los estudios, se comprobó que la mutación inactivaba la enzima y presentaba entre un 3% -4% de actividad residual (14).
Pese a la existencia de reportes de mutaciones, en algunas de las formas infantiles presentadas con afectación multisistémica, no está completamente dilucidado el mecanismo molecular por el que la enfermedad ocurre, ya que no se han detectados mutaciones en el gen de la FFQ muscular (13).
POTENCIAL DE LA BIOINFORMÁTICA PARA EL ESTUDIO DE LA ENFERMEDAD
Con el gran número de algoritmos y modelos matemático-computacionales disponibles para el estudio de las moléculas, la bioinformática se perfila como un elemento clave para el entendimiento de las enfermedades en las que no existe una clara asociación entre el genotipo y el fenotipo, como lo es la enfermedad de Tarui. En este sentido, dos áreas específicas de la bioinformática son de interés: la modelación de estructuras y la biología de sistemas.
Respecto al análisis de secuencias, un elemento básico para el análisis de estructuras, la caracterización de las FFQ humanas ha permitido determinar que las FFQ muscular y hepática comparten 68,6% de identidad de secuencia de aminoácidos, mientras que la muscular y la plaquetaria un 70,3%, y la hepática y la plaquetaria comparten el 66,6% (3,7). Además, modelos estructurales 3D de las isoformas plaquetaria y muscular han sido recientemente determinados. La primera estructura del tetrámero de la FFQ de la isoforma de plaquetas humanas, en complejo con ATP-Mg21 y ADP, se publicó en 2015 (1,9). Un año antes, Kloos y colaboradores (2014), lograron la cristalización y un análisis preliminar de la FFQ muscular humana, en el que verificaron que la enzima se disocia en un dímero (3,8). Estos avances brindan la oportunidad de nuevas alternativas para entender la regulación alostérica y las bases moleculares de los mecanismos asociados a los activadores o inhibidores de la función de la enzima. Además, en la enfermedad de Tarui vista como glucogenosis primaria, se lograría la evaluación y modelado del efecto de las mutaciones de la enzima y cómo contribuyen a la pérdida de su actividad. Adicionalmente, dado que las mutaciones en la FFQ también han sido asociadas a otros tipos de padecimientos como diabetes y malignidad, estos posibles estudios no se limitarían únicamente a la enfermedad de Tarui, sino que en general permitirían una mejor comprensión de todos los mecanismos biológicos en los que la enzima participa.
Como ha sido verificado en diversos casos reportados, ya sea por la existencia de las mismas mutaciones en la FFQ con diferente sintomatología entre pacientes, o bien, en el caso de la enfermedad vista como glucogenosis secundaria, el entendimiento de la enfermedad de Tarui es de alta complejidad. Ofrece un desafío para su estudio que comprende mecanismos regulatorios que no podrían ser explicados únicamente con el estudio de la estructura tridimensional de forma aislada. Es por ello que una segunda área de la bioinformática de interés para el estudio de esta EAG es la biología de sistemas, con la cual se evalúan propiedades que únicamente pueden emerger a partir del análisis de los sistemas biológicos vistos como un todo y no como elementos individuales.
Un ejemplo del potencial de la biología de sistemas puede ser considerado con el modelo matemático presentado por Lambeth y colaboradores, el cual representa dinámicamente la vía de la glicólisis hasta la producción de energía y lactato en el músculo esquelético de mamíferos. El modelo se construyó con parámetros cinéticos obtenidos a partir de la literatura y con restricciones estequiométricas, balance de masa y con una termodinámica basada en la ley de acción de masas combinada. La topología general es presentada en la Figura 1 (los números en los círculos rojos indica un paso enzimático en el orden usual de la glicólisis). Los mismos autores mencionan que el principal resultado del análisis fue el acoplamiento de la vía con la red de regulación de la ATPasa, haciendo que el sistema en estado estable sea más sensible a la actividad de ATPasa externa y no a mecanismos internos de la vía, lo cual permite la interconexión con otras vías y explica cómo los cambios en la glicólisis logran tener un efecto en vías alternativas para modular la cinética del ATP en el músculo esquelético (39).
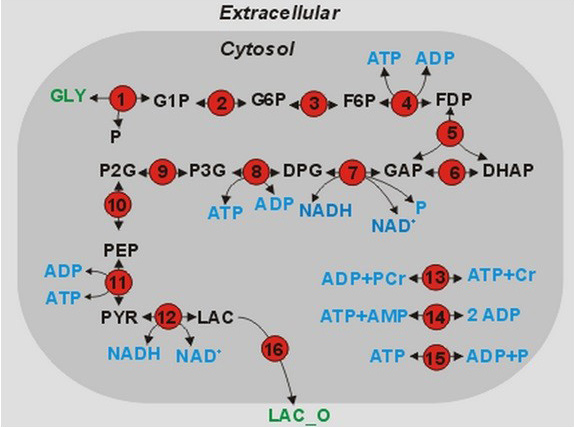
Figura 1. Topología del modelo de la glicosis en músculo esquelético. (Imagen del reposiorio JWS Online /http://jjj.biochem.sun.ac.za/) del trabajo de Lambeth & Kushmerick, 2002).
Con este modelo es posible realizar simulaciones en línea, incluyendo la enfermedad de Tarui (http://jjj.biochem.sun.ac.za/models/?id=lambeth). Inicialmente, utilizando el sistema de ecuaciones diferenciales planteado, la constante de reacción del paso catalizado por la FFQ se redujo a la mitad (Vfpfk, de 0.056 a 0.028), con el objetivo de valorar el efecto en la producción de los metabolitos finales del modelos, NADH (el valor bajó a un 9% del normal) y piruvato (el valor bajó a un 60% del normal). Estos dos metabolitos son particularmente relevantes en el contexto de la fisiopatología de la enfermedad, pues el bloqueo de la glicólisis impide la producción de éstos y su continuidad a rutas metabólicas (aeróbicas o anaeróbicas), lo cual compromete la disponibilidad de energía en los tejidos, principalmente músculo. Si existiesen otras alternativas para modular la producción, podría usarse como estrategia para solventar la necesidad energética ante la deficiencia de la FFQ, y justamente las predicciones in silico del modelo matemático propone alternativas para ello. Al realizar el análisis de sensibilidad en el modelo de ecuaciones diferenciales y con el objetivo de determinar los puntos del modelo que permitan aumentar los valores de dichos productos (ver tabla 3), se obtuvo que los pasos del modelo donde actúan la piruvato quinasa (PQ, paso V11) y la deshidrogenasa láctica (DHL, paso V12) fueron los más representativos para reponer los valores del NADH y piruvato.
En la tabla 3, los valores extremos positivos de los coeficientes de control (rojo, paso V1) indican que la producción de NADH y piruvato se favorece al incrementar la reacción asociada, mientras que los valores extremos negativos (verde) son lo que más consumen dichas moléculas. Aunque algebraicamente aumentar los valores del paso V1 significaría un aumento de los productos, el bloqueo del paso de la FFQ no tendría efecto. Así, la inhibición de las reacciones V11 y V12 supondría la disminución del consumo de NADH y piruvato. En la simulación, la inhibición al 50% de cada una de las actividades en la respectiva constante de reacción, muestra un aumento de NADH al 70% del normal (enfermo era 9%) y un 95% de piruvato respecto al normal (enfermo era 60%).

Tabla 3. Coeficientes de control para NADH y piruvato en el modelo matemático.
Fuente: Repositorio JWS Online (http://jjj.biochem.sun.ac.za/), Lambeth & Kushmerick, 2002 (39)
De forma acoplada a otras vías de regulación, esto indicaría que puede modularse la actividad o la cinética asociada a las enzimas piruvato quinasa y deshidrogenasa láctica para reponer la producción de energía de forma alternativa para contrarrestar los efectos en la enfermedad de Tarui. En este sentido, aspectos como el redireccionamiento de la fuente de energía hacia los lípidos o el flujo de carbono a través de la vía de las pentosas fosfato (1)(9), serían aspectos que podrían estudiarse como un sistema metabólico que, en conjunto con la FFQ, podrían brindar explicaciones referentes a la sintomatología o de opciones para posibles tratamientos.
Además, en otras posibles líneas de modulación, se ha reportado que la activación y la inhibición alostérica de la enzima por más de diez metabolitos y en respuesta a la señalización hormonal resultan en la regulación del flujo glucolítico para satisfacer las necesidades de energía de las células (19). Estos reguladores incluyen ATP, lactato, AMP, cAMP, ADP, fructosa-2,6-bisfosfato, citrato y acil-CoA (35) y se ha demostrado que estos moduladores alostéricos ejercen propiedades estimulantes o inhibitorios que interfieren con el equilibrio entre dímeros y tetrámeros de la enzima, lo cual hace parecer plausible que moléculas como el ATP, el citrato y el lactato inhiban la FFQ mediante la estabilización de su conformación dimérica, mientras que ADP, AMP, cAMP y fructosa-2,6-bisfosfato favorecerían la formación de tetrámeros (37).
Adicionalmente, a un nivel génico (microARNs que regulen la expresión del gen de la FFQ) o bien con modificaciones post traduccionales de la misma enzima u otras proteínas y hormonas, podrían ser claves en la regulación de la actividad de la enzima (1). Por ejemplo, se ha comprobado que la fosforilación de la FFQ resulta de la estimulación por hormonas tales como epinefrina, insulina o serotonina en el músculo esquelético, y todas estas hormonas tienen la capacidad de activar glucólisis (37). Así, es posible apreciar la complejidad metabólica en la que la FFQ está inmersa, dado que está involucrada en diversas vías que terminan por regular su actividad o en aquellas que ella misma regula. Toda esa variedad podría explicar la poca claridad entre el genotipo-fenotipo en la enfermedad de Tarui.
CONCLUSIONES
Se ha presentado una descripción de la enfermedad de Tarui, un padecimiento del almacenamiento del glucógeno que afecta el músculo esquelético y ha sido reportada en al menos 100 individuos. Es ocasionada por la deficiencia de la actividad fosfofructoquinasa y se hereda con un patrón autosómico recesivo. Dado que esta enzima participa en la glicólisis y la regula, su afectación compromete la producción de energía a partir del consumo de la glucosa, con síntomas variables, pero normalmente incluye afectación muscular con intolerancia al ejercicio, mialgias, debilidad y fatiga, así como cuadros de hemólisis. Dado que varias de las glucogenosis podrían presentar síntomas similares, el diagnóstico de la enfermedad se basa en el análisis de pruebas de laboratorio como la biopsia de músculo, pruebas inmunohistoquímicas, medición de la actividad de la enzima y prueba de glucosa exógena previo a ejercicio. Además, se han detectado diferentes mutaciones de la enzima, pero las variaciones en la presentación clínica no permiten establecer una relación genotipo-fenotipo clara, por lo que se ha atribuido dicha variación a los diversos mecanismos regulatorios en los que la fosfofructoquinasa participa. Esto brinda posibles opciones de trabajo futuro referente al entendimiento de la biología de la enfermedad, en las que herramientas matemático-computacionales de la bioinformática brindarían alternativas de análisis tanto de la estructura de la enzima como de las redes metabólicas de forma entrecruzada. Esto permitiría mejorar las opciones diagnósticas, pronósticas y, especialmente, de tratamiento, el cual no existe hasta el momento.
REFERENCIAS
- Kilimann MW, Oldfors A. Glycogen pathways in disease: new developments in a classical field of medical genetics. J Inherit Metab Dis. 2015;38(3):483-487. doi:10.1007/s10545-014-9785-5.
- Berardo A, DiMauro S, Hirano M. A diagnostic algorithm for metabolic myopathies. Curr Neurol Neurosci Rep. 2010;10(2):118-126. doi:10.1007/s11910-010-0096-4.
- Gazzerro E, Andreu AL, Bruno C. Neuromuscular disorders of glycogen metabolism. Curr Neurol Neurosci Rep. 2013;13(3):333. doi:10.1007/s11910-012-0333-0.
- Volpi L, Ricci G, Orsucci D, et al. Metabolic myopathies: functional evaluation by different exercise testing approaches. Musculoskelet Surg. 2011;95(2):59-67. doi:10.1007/s12306-011-0096-9.
- Chalès G, Coiffier G, Guggenbuhl P. Manifestations ostéoarticulaires des mucopolysaccharidoses et des glycogénoses. Rev du Rhum Monogr. 2011;78(4):254-261. doi:10.1016/j.monrhu.2011.03.008.
- Özen H. Glycogen storage diseases : New perspectives. 2007;13(18):2541-2553.
- Hicks J, Wartchow E, Mierau G. Glycogen storage diseases: a brief review and update on clinical features, genetic abnormalities, pathologic features, and treatment. Ultrastruct Pathol. 2011;35(5):183-196. doi:10.3109/01913123.2011.601404.
- Fujii H, Miwa S. Other erythrocyte enzyme deficiencies associated with non-haematological symptoms: phosphoglycerate kinase and phosphofructokinase deficiency. Best Pract Res Clin Haematol. 2000;13(1):141-148. doi:10.1053/beha.1999.0062.
- Musumeci O, Bruno C, Mongini T, et al. Clinical features and new molecular findings in muscle phosphofructokinase deficiency (GSD type VII). Neuromuscul Disord. 2012;22(4):325-330. doi:10.1016/j.nmd.2011.10.022.
- Tarui S, Giichi O, Ikura Y, Tanaka T, Suda M, Nishikawa M. Phosphofructokinase deficiency in skeletal muscle. A new type of glycogenosis. Biochem Biophys Res Commun. 1965;19(4):517-523. doi:10.1016/0006-291X(65)90156-7.
- Ronquist G, Waldenstro A. Imbalance of plasma membrane ion leak and pump relationship as a new aetiological basis of certain disease states. 2003:517-526.
- Dimauro S, Garone C. Metabolic disorders of fetal life: glycogenoses and mitochondrial defects of the mitochondrial respiratory chain. Semin Fetal Neonatal Med. 2011;16(4):181-189. doi:10.1016/j.siny.2011.04.010.
- Malfatti E, Birouk N, Romero NB, et al. Juvenile-onset permanent weakness in muscle phosphofructokinase deficiency. J Neurol Sci. 2012;316(1-2):173-177. doi:10.1016/j.jns.2012.01.027.
- Auranen M, Palmio J, Ylikallio E, et al. PFKM gene defect and glycogen storage disease GSDVII with misleading enzyme histochemistry. Neurol Genet. 2015;1(1):e7-e7. doi:10.1212/NXG.0000000000000007.
- Mcardle B. Muscle glycogenoses : an overview. 2007;(i):35-41.
- Kinase P, Mutase P. Tarui disease and distal glycogenoses : clinical and genetic update. 2007;(4):105-107.
- Ono A, Kuwajima M, Kono N, et al. Glucose infusion paradoxically accelerates degradation of adenine nucleotide in working muscle of patients with glycogen storage disease type VII. 1995;(January).
- Massa R, Sancesario G, Bernardi G. Muscle phosphofructokinase deficiency. Neurology. 1997;49(3):899. http://www.ncbi.nlm.nih.gov/pubmed/9305371. Accedido enero 28, 2016.
- Webb B a., Forouhar F, Szu F-E, Seetharaman J, Tong L, Barber DL. Structures of human phosphofructokinase-1 and atomic basis of cancer-associated mutations. Nature. 2015;523(7558):111-114. doi:10.1038/nature14405.
- Ronquist G, Waldenstro A. Increased erythrocyte content of Ca 2 1 in patients with Tarui ’ s disease. 2001:97-102.
- Finsterer J, Stöllberger C, Kopsa W. Neurologic and cardiac progression of glycogenosis type VII over an eight-year period. South Med J. 2002;95(12):1436-1440. http://www.ncbi.nlm.nih.gov/pubmed/12597313. Accedido enero 28, 2016.
- Nakajima H, Raben N, Hamaguchi T, Yamasaki T. Phosphofructokinase Deficiency Past, Present and Future. Curr Mol Med. 2002;2(2):197-212. doi:10.2174/1566524024605734.
- Finsterer J, Stöllberger C. Progressive mitral valve thickening and progressive muscle cramps as manifestations of glycogenosis VII (Tarui’s Disease). Cardiology. 2008;110(4):238-240. doi:10.1159/000112406.
- Nichols RC, Rudolphi O, Ek B, Exelbert R, Plotz PH, Raben N. Glycogenosis Type VII ( Tarui Disease ) in a Swedish Family : Two Novel Mutations in Muscle Phosphofructokinase Gene ( PFK-M ) Resulting in Intron Retentions. 1996:59-65.
- Hays AP, Hallett M, Delfs J, et al. Muscle phosphofructokinase deficiency: Abnormal polysaccharide in a case or late-onset myopathy. Neurology. 1981;31(9):1077-1077. doi:10.1212/WNL.31.9.1077.
- Haller R, Vissing J. No spontaneous second wind in muscle phosphofructokinase deficiency. Neurology. 2004;62(1):82-86. http://www.ncbi.nlm.nih.gov/pubmed/14718702. Accedido enero 21, 2016.
- Di Mauro S. Muscle glycogenoses: an overview. Acta Myol. 2007;26(1):35-41. http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2949320&tool=pmcentrez&rendertype=abstract. Accedido enero 28, 2016.
- Haller R, Lewis S. Glucose-induced exertional fatigue in muscle phosphofructokinase deficiency. N Engl J Med. 1991;324(6):364-369. doi:10.1056/NEJM199102073240603.
- Sun B, Brooks E, Koeberl D. Preclinical Development of New Therapy for Glycogen Storage Diseases. Curr Gene Ther. 2015;15(4):338-347. doi:10.2174/1566523215666150630132253.
- Tsujino S, Servidei S, Tonin P, Shanske S, Azan G, DiMauro S. Identification of three novel mutations in non-Ashkenazi Italian patients with muscle phosphofructokinase deficiency. Am J Hum Genet. 1994;54(5):812-819. http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1918246&tool=pmcentrez&rendertype=abstract. Accedido enero 21, 2016.
- Davidson M, Miranda AF, Bender AN, DiMauro S, Vora S. Muscle phosphofructokinase deficiency. Biochemical and immunological studies of phosphofructokinase isozymes in muscle culture. J Clin Invest. 1983;72(2):545-550. doi:10.1172/JCI111002.
- Nakajima H, Kono N, Yamasaki T, et al. Genetic defect in muscle phosphofructokinase deficiency. Abnormal splicing of the muscle phosphofructokinase gene due to a point mutation at the 5’-splice site. J Biol Chem. 1990;265(16):9392-9395. http://www.ncbi.nlm.nih.gov/pubmed/2140573. Accedido enero 28, 2016.
- Vives-Corrons J-L, Koralkova P, Grau JM, Mañú Pereira MDM, Van Wijk R. First description of phosphofructokinase deficiency in spain: identification of a novel homozygous missense mutation in the PFKM gene. Front Physiol. 2013;4(December):393. doi:10.3389/fphys.2013.00393.
- Sherman JB, Raben N, Nicastri C, et al. Common mutations in the phosphofructokinase-M gene in Ashkenazi Jewish patients with glycogenesis VII–and their population frequency. Am J Hum Genet. 1994;55(2):305-313. http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1918380&tool=pmcentrez&rendertype=abstract. Accedido enero 21, 2016.
- Brüser A, Kirchberger J, Schöneberg T. Altered allosteric regulation of muscle 6-phosphofructokinase causes Tarui disease. Biochem Biophys Res Commun. 2012;427(1):133-137. doi:10.1016/j.bbrc.2012.09.024.
- Raben N, Sherman JB. Mutations in muscle phosphofructokinase gene. Hum Mutat. 1995;6(1):1-6. doi:10.1002/humu.1380060102.
- Sola-Penna M, Da Silva D, Coelho WS, Marinho-Carvalho MM, Zancan P. Regulation of mammalian muscle type 6-phosphofructo-1-kinase and its implication for the control of the metabolism. IUBMB Life. 2010;62(11):791-796. doi:10.1002/iub.393.
- Kloos M, Bruser A, Kirchberger J, Schoneberg T, Strater N. Crystallization and preliminary crystallographic analysis of human muscle phosphofructokinase, the main regulator of glycolysis. Acta Crystallogr F Struct Biol Commun. 2014;70:578-582. doi:24817713.
- Lambeth MJ, Kushmerick MJ. A Computational Model for Glycogenolysis in Skeletal Muscle. Ann Biomed Eng. 2002;30(6):808-827. doi:10.1114/1.1492813.
INFORMACIÓN DEL ARTÍCULO
Autor para la correspondencia: Jose Arturo Molina Mora. Centro de Investigaciones en Enfermedades Tropicales, Universidad de Costa Rica. E-mail: jose.molinamora@ucr.ac.cr





